Эволюционная
Теория
Пола:
Повышенная смертность мужского пола
и тип гаметности
 Среди многих непонятных фактов,
связанных с дифференциацией полов (адаптивных корреляций
вторичного соотношения полов, высоких значений первичного
и др.), повышенная смертность мужского пола как
общебиологическое явление, тесно связанное с эволюционными
преимуществами раздельнополости, занимает особое место.
Исследуем ли мы различные виды (человек, животные или
растения), разные уровни организации (организм, орган, ткань
или клетка) или устойчивость к разным вредным факторам среды
(низкие и высокие температуры, голод, яды, паразиты, болезни
и др.) — везде и всюду одна и та же картина: повышенная
смертность или повреждаемость мужских систем по сравнению
с соответствующими женскими. Среди многих непонятных фактов,
связанных с дифференциацией полов (адаптивных корреляций
вторичного соотношения полов, высоких значений первичного
и др.), повышенная смертность мужского пола как
общебиологическое явление, тесно связанное с эволюционными
преимуществами раздельнополости, занимает особое место.
Исследуем ли мы различные виды (человек, животные или
растения), разные уровни организации (организм, орган, ткань
или клетка) или устойчивость к разным вредным факторам среды
(низкие и высокие температуры, голод, яды, паразиты, болезни
и др.) — везде и всюду одна и та же картина: повышенная
смертность или повреждаемость мужских систем по сравнению
с соответствующими женскими.
Гамильтон приводит обзор
дифференциальной смертности полов для 70 видов, включая
такие различные формы жизни, как нематоды, моллюски,
ракообразные, насекомые, паукообразные, птицы, рептилии,
рыбы, млекопитающие. Согласно этим данным, у 62 видов (89%)
средняя длительность жизни самцов короче, чем у самок;
у большинства остальных нет разницы, и лишь в отдельных
случаях длительность жизни самцов больше, чем у самок.
По данным ООН, за небольшими
исключениями, во всех странах мира отмечена более высокая
средняя продолжительность жизни женщин. У человека
повышенная смертность мужского пола наблюдается почти по
всем болезням, кроме коклюша и некоторых гонококковых
инфекций. Аналогичная картина наблюдается не только
у животных и человека, но и у растений. Следовательно,
повышенная смертность мужского пола — явление универсальное.
Существующие
теории не в состоянии
удовлетворительно объяснить это явление.
Эволюционная теория
пола рассматривает повышенную смертность мужского пола как
выгодную для популяции форму информационного контакта со
средой, осуществляемую через элиминацию вредным фактором
среды части особей популяции. Изображая кривую смертности
популяции от того или иного вредного фактора среды, мы тем
самым получаем кривую фенотипического распределения
популяции по соответствующему признаку. Следовательно, чем
больше дисперсия кривой смертности, тем больше дисперсия
кривой фенотипического распределения и тем чувствительнее
популяция к данному вредному фактору среды. Таким образом,
если первыми жертвами вредного фактора среды являются
мужские особи, то это означает, что фенотипическая
дисперсия у мужского пола больше, чем у женского.
Значит, более тесная связь мужского пола со средой
обеспечивается большей величиной его фенотипической
дисперсии по сравнению с женским полом.
Тип гаметности и
пол
Конфликт по
консервативной и оперативной задачам эволюции лежит в основе
не только возникновения дифференциации полов, но и вообще
любой дифференциации. В связи с полом такой конфликт
возникал по крайней мере дважды. Первый раз, когда
существовала изогамия на уровне клеток, возникли конфликтные
требования к их размерам. В оперативную тенденцию входила
задача найти другую клетку, для чего необходимы подвижность
и малые размеры, облегчающие решение этой задачи.
В консервативную тенденцию входили задачи сохранения
образовавшейся зиготы, обеспечения ее материальными
и энергетическими ресурсами, защитными оболочками и др., что
связано с большими размерами. Результат разрешения этого
конфликта — появление дифференциации гамет по размеру
и подвижности на яйцеклетки и сперматозоиды.
Та же самая дилемма
возникла при дифференциации по типу гаметности.
Необходимо было иметь возможность пробовать рецессивные гены
прежде, чем дать им закрепиться, что можно было делать
только в их гомозиготном состоянии в гетерогаметном наборе
хромосом (XY). Это оперативная тенденция, тогда как
гетерозиготная комбинация генов в гемигаметном наборе (XX),
при которой ущербный рецессивный ген не проявляется,
реализует консервативную тенденцию.
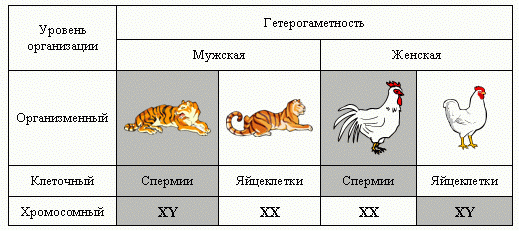
В процессе эволюции
у большинства видов обе оперативные подсистемы (малые гаметы
и гетерогаметная конституция XY) оказались у мужского пола,
а обе консервативные подсистемы (крупные гаметы
и гемигаметная конституция XX) — женского. Это виды
с гаметностью типа Drosophila
(Таблица).
Таблица.
Консервативные и оперативные подсистемы на разных
уровнях организации у млекопитающих и птиц.
Однако у некоторых
видов, в их эволюции, направления этих двух дифференциации
не совпали. У женского пола оказались консервативные
яйцеклетки в сочетании с оперативной гетерогаметностью, а у
мужского — сочетание оперативных сперматозоидов
с консервативной гемигаметностью. Это виды с гаметностью
типа Abraxas (птицы, бабочки, ручейники, некоторые
рыбы и др.).
Следовательно, система
гаметности типа Drosophila последовательна, а система
типа Abraxas противоречива. Этим, возможно,
объясняется тот факт, что видов типа Drosophila
несравненно больше, чем типа Abraxas, если считать,
что выбор типа происходил чисто случайно (независимо от
пола).
Влияние типа
гаметности и полигамии
Эффект
дифференциальной смертности полов состоит из двух
компонентов (Рис.
1).
Первый—это вклад типа гаметности, который хорошо объясняется
теорией дисбаланса генов. Второй компонент—вклад от
специализации полов на уровне популяции. Поскольку
популяционный эффект возникает только в панмиктной, свободно
скрещивающейся популяции и связан, видимо, со степенью
полигамии, то при объяснении наблюдаемой картины различной
смертности полов необходимо учесть как тип гаметности, так и
моно- или полигамность вида.
У моногамов
популяционные эффекты сведены к минимуму (у строгих
моногамов к нулю) и повышенная смертность наблюдается у
гетерогаметного пола (у самцов типа Drosophila
и самок типа Abraxas). Факты
удовлетворительно объясняются теорией дисбаланса генов. У
полигамных видов типа Drosophila
на эффект от гаметности накладывается еще и популяционный
эффект, усиливая его; этим объясняется отмеченная
максимальная разница у таких видов. Для полигамных видов
типа Abraxas популяционный
эффект направлен против эффекта гаметности: гетерогаметность
приводит к сокращению длительности жизни самок, а
популяционный механизм—самцов. Поэтому можно ожидать, что
разница в смертности у них будет менее выраженной, чем для
полигамов типа Drosophila, при
одинаковой степени полигамии. Из сказанного следует, что у
моногамных видов типа Abraxas
смертность должна быть выше у самок, что в действительности
и наблюдается .
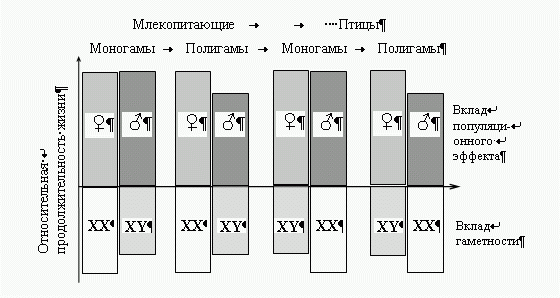
Рис.
1
Вклад
специализации полов на уровне популяции (верхняя часть
рисунка) и вклад типа гаметности (нижняя
часть рисунка) в дифференциальную смертность полов.
Популяционный эффект возникает только в свободно
скрещивающейся (полигамной) популяции, а у моногамов он
практически не проявляется. Для полигамов
характерна повышенная смертность мужских особей. Мужской пол
и гетерогаметная конституция (XY) выполняют
оперативную задачу, женский пол и гомогаметная конституция
(XX)—консервативную. У некоторых видов
направления этих двух дифференциаций не совпадают (например
у птиц). Таким образом у полигамных видов с
женской гетерогаметностью популяционный эффект направлен
против эффекта гаметности, а у полигамов с
мужской гетерогаметностью эти эффекты совпадают.
Таким образом, новый
подход хорошо объясняет наблюдаемую картину смертности
полов. Строго говоря, для реализации популяционных
механизмов специализации полов важно, чтобы смертность
мужского пола “опережала” смертность женского. Во взрослых
возрастах также мужская смертность превышает женскую.
Отчасти это может быть связано с тем, что все “новые”
болезни, болезни “века” или “цивилизации” (инфаркт,
атеросклероз, гипертония, шизофрения и др.), как правило,
болезни мужского пола.
Существующие теории—хромосомного
дисбаланса и метаболическая—рассматривают
дифференциальную смертность полов как пассивное следствие
хромосомной конституции или уровня метаболизма. Подробнее
Новая теория рассматривает повышенную смертность мужского
пола как активное средство, повышающее эволюционную
устойчивость популяции и вскрывает эволюционный смысл,
логику явления дифференциальной смертности, связывая его с
разной нормой реакции полов.
Длительность жизни,
генотип и среда
►
Подробнее о
дифференциальной смертности полов
Первая публикация:
Дифференциальная смертность полов и норма
реакции.
Геодакян В. А. Биол. журн. Армении, 1973, т. 26, № 6,
с. 3–11.
Популярное изложение:
Женщины живут дольше?
(А. Петров). Труд, 2 Августа 1983 г., с. 3.
Ева овдовела первой? (О. Парфенова). Труд-7,
29 Марта 2001 г., с. 22.
Более полное научное
изложение:
Эволюционная логика
дифференциации полов и долголетие.
Геодакян В. А. Природа, 1983б, № 1, с. 70–80.
Наиболее свежая
публикация:
Теория дифференциации
полов в проблемах человека.
Геодакян В. А. Человек в системе наук. М., Наука,
1989, с. 171–189.
|