Эволюционная
Теория
Пола:
Повышенная смертность мужского пола
(2): Зависимость
смертности популяции от генотипа и среды
 Длительность жизни,
как и любой признак, определяется генотипом и средой.
Представим следующие идеализированные ситуации. Поместим
мысленно клон генотипически идентичных особей популяции в
оптимальную среду. Тогда длительность жизни всех особей
будет одинакова и кривая смертности будет иметь
прямоугольную форму (дисперсия σ = 0
Рис. 2А).
Теперь вместо клона возьмем гетерогенную популяцию в
оптимальной среде, позволяющей реализоваться полностью всем
генотипам—появится некий разброс в длительности жизни,
обусловленный генотипическим разнообразием (дисперсия σген
Рис. 2Б).
Вновь вернемся к клону, но поместим его в реальную среду.
Появится некий разброс длительности жизни, обусловленный
только средой (дисперсия σср
Рис. 2В).
Поместим генотипически гетерогенную популяцию в реальную
среду. Тогда суммарная дисперсия будет включать как
генотипическую, так и средовую компоненты (σ = σген
+ σср
Рис. 2Г).
Наконец представим себе ситуацию, когда смертность
определяется чисто стохастически, только случаем (то есть
средой). Тогда генотип не будет играть никакой роли в
определении длительности жизни и последняя не будет зависеть
ни от возраста, ни от здоровья. В этом случае кривая
смертности будет иметь экспоненциальную форму (Рис.
2Д). Длительность жизни,
как и любой признак, определяется генотипом и средой.
Представим следующие идеализированные ситуации. Поместим
мысленно клон генотипически идентичных особей популяции в
оптимальную среду. Тогда длительность жизни всех особей
будет одинакова и кривая смертности будет иметь
прямоугольную форму (дисперсия σ = 0
Рис. 2А).
Теперь вместо клона возьмем гетерогенную популяцию в
оптимальной среде, позволяющей реализоваться полностью всем
генотипам—появится некий разброс в длительности жизни,
обусловленный генотипическим разнообразием (дисперсия σген
Рис. 2Б).
Вновь вернемся к клону, но поместим его в реальную среду.
Появится некий разброс длительности жизни, обусловленный
только средой (дисперсия σср
Рис. 2В).
Поместим генотипически гетерогенную популяцию в реальную
среду. Тогда суммарная дисперсия будет включать как
генотипическую, так и средовую компоненты (σ = σген
+ σср
Рис. 2Г).
Наконец представим себе ситуацию, когда смертность
определяется чисто стохастически, только случаем (то есть
средой). Тогда генотип не будет играть никакой роли в
определении длительности жизни и последняя не будет зависеть
ни от возраста, ни от здоровья. В этом случае кривая
смертности будет иметь экспоненциальную форму (Рис.
2Д).
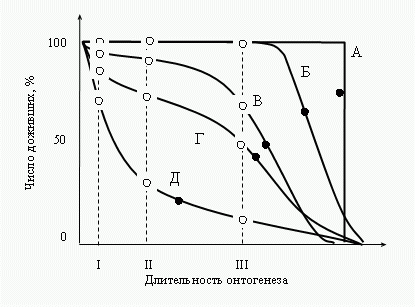   Рис.
2 Рис.
2
Зависимость
смертности популяции от генотипа и среды.
А—для клона в идеальной среде (σген
= 0, σср
= 0); Б—для гетерогенной популяции в идеальной среде (σген
≠ 0, σср
= 0);
В—для клона в реальной среде (σген
= 0, σср
≠ 0); Г—для гетерогенной популяции в реальной среде (σген
≠ 0, σср
≠ 0);
Д—смертность определяется чисто стохастически.
I—средняя
длительность утробной жизни;
II—средний
возраст появления репродуктивной функции,
III—средний
возраст
утраты репродуктивной функции, ● —средняя продолжительность
жизни, ○ —средние длительности стадий онтогенеза.
Кривые Б, В и Г как и
все реальные случаи, встречающиеся в жизни, представляют
собой промежуточные переходы между крайними типами А
(смертность определяется полностью генотипом) и Д
(смертность зависит только от среды). Следовательно, по мере
“эмансипации” популяции от среды картина ее смертности будет
приближаться к прямоугольному типу А и удаляться от
экспоненциального типа Д. Значит, чем оптимальнее среда, тем
картина смертности популяции ближе к типу А, чем
экстремальнее—тем ближе к типу Д.
При переходе
популяции из оптимальной среды в экстремальную, можно
видеть, что смертность сильно влияет на среднюю
продолжительность жизни и пострепродуктивного периода и
фактически не сказывается на средней длительности периодов
утробного развития, роста и репродукции (Рис.
2).
Это значит, что в экстремальных условиях среды уменьшение
средней продолжительности жизни идет за счет средовой
компоненты смертности, а не генотипической. Таким образом,
применение правила полового диморфизма дает основание
предположить, что генотипическая средняя длительность жизни
мужчин должна быть больше, чем женщин. И если можно было бы
полностью устранить влияние среды то мужчины жили бы дольше
женщин.
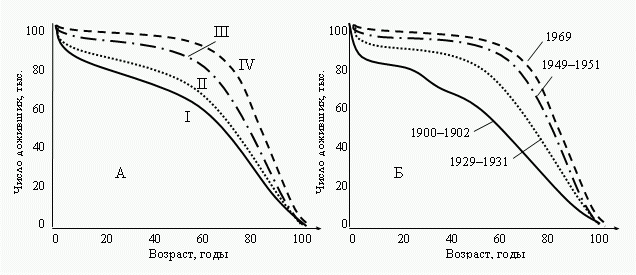
Рис. 3
Кривые
продолжительности жизни для населения США. А: Число выживших
на 100 000 рожденных
(США,
1939–1941 гг.): I—негры мужчины; II—негры женщины;
III—белые мужчины; IV—белые
женщины.
Б: продолжительность жизни населения США в разные годы
(Комфорт, 1967).
Таковы выводы,
которые можно сделать из теории. Рассмотрим известные факты.
Если сравнить друг с другом в разной степени
“эмансипированные” от среды группы, то увидим, что характер
кривой смертности закономерно меняется в указанном теорией
направлении (Рис.
3А).
Чем более “эмансипирована” группа от среды, тем ближе ее
кривая смертности к “прямоугольному” типу, и наоборот.
Бóльшая “эмансипация” женского пола по сравнению с мужским
обусловлена его более широкой нормой реакции; белого
населения США по сравнению с неграми, а также современной
популяции по сравнению с популяциями прошлых лет обусловлена
социально-экономическими причинами (питание, медицинское
обслуживание) (Рис.
3Б).
Таким образом,
предлагается гипотеза, которая снимает противоречия между
известными корреляциями и правилом полового диморфизма. Она
объясняет, почему средняя длительность жизни больше у
женщин, а "чемпионы"-долгожители—мужчины. Например, в
центральной части Южного Закавказья в 40-х годах прошлого
века из 15 долгожителей в возрасте 110–140 лет 14 были
мужчины. В Абхазии в 20-х годах XX
века все верхние ступени возрастной лестницы также занимали
мужчины [14]. Она позволяет понять и тот, казалось бы,
парадоксальный факт, что феномен долгожительства встречается
в популяциях, живущих отнюдь не в оптимальных условиях
[3,10].
Если феномен
долгожительства действительно обусловлен повышением
дисперсии длительности жизни в экстремальных условиях среды,
тогда он должен быть тесно связан со стрессом как
передатчиком экологической информации у животных, должен
сопровождаться повышенной смертностью (в первую очередь
детской), повышенной дисперсией других признаков, повышенным
вторичным соотношением полов (рождаемостью мальчиков) и
ростом полового диморфизма. Популяции, в которых встречается
феномен долголетия, должны иметь более близкую к
экспоненциальному типу кривую смертности, характерную для
экстремальных условий среды, в то время как кривая
смертности популяции в оптимальной среде должна быть ближе к
прямоугольному типу, при котором феномен долголетия не
наблюдается.
[
3]. Комфорт А.
Биология старения. М.: Мир, 1967, с. 400.
[10].
Давыдовский И.В. Геронтология. М.: Медицина, 1966, с.
300.
[14].
Архипов И. - Вокруг света, 1978, № 10, с. 60-62.
Подробнее о
дифференциальной смертности полов
Первая публикация:
Дифференциальная смертность полов и норма
реакции.
Геодакян В. А. Биол. журн. Армении, 1973, т. 26, № 6,
с. 3–11.
Популярное изложение:
Женщины живут дольше?
(А. Петров). Труд, 2 Августа 1983 г., с. 3.
Ева овдовела первой? (О. Парфенова). Труд-7,
29 Марта 2001 г., с. 22.
Более полное научное
изложение:
Эволюционная логика
дифференциации полов и долголетие.
Геодакян В. А. Природа, 1983б, № 1, с. 70–80.
Наиболее свежая
публикация:
Теория дифференциации
полов в проблемах человека.
Геодакян В. А. Человек в системе наук. М., Наука,
1989, с. 171–189.
|